|
|
Post by kammy on Sept 14, 2009 21:55:30 GMT -5
I've been studying how are some of these processes are done in the labs. There's a site called www.jove.com that costs money to have a subscription, but, they give you a limited free day. I signed up for the free 1 day subscription. I entered the search word 'baculovirus' and got to see how they microinject the caterpillars with the polyhedron... most impressive...  And, here's a video on Pubmed that hosts some of Jove's, this one is called: Wolbachia Bacterial Infection in Drosophila www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2532937"Journal of Visualized Experiments (JoVE) is an online research journal employing visualization to increase reproducibility and transparency in biological sciences." |
|
|
|
Post by kammy on Sept 16, 2009 6:32:30 GMT -5
|
|
|
|
Post by Jill on Sept 16, 2009 8:14:56 GMT -5
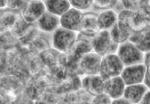
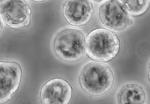
Kammy, I think we may be looking at the same things... The images you have posted are Infected and uninfected SF9 cell. SF 9 insect cells are just that- from insects or worms The source, vaccines and Chemotherapy- cancer treatment. Baculovirus is used in Cancer treatment. The SF9 cells- are the most susceptible to the baculovirus- Search Bing or Google-there are over 252,000 hits- Baculovirus + patents & or vaccines Per the Protein Data Bank: www.rcsb.org/pdb/results/results.do?gotopage=5 SF 9 cells yields 5 pages - of hits (note: Bovine) Here is one- at random: www.rcsb.org/pdb/explore.do?structureId=1INP CRYSTAL STRUCTURE OF INOSITOL POLYPHOSPHATE 1-PHOSPHATASE AT 2.3 ANGSTROMS RESOLUTION York, J.D., Ponder, J.W., Chen, Z.W., Mathews, F.S., Majerus, P.W. (1994) Biochemistry 33: 13164-13171 PubMed: 7947723 View PubMed Abstract at NCBI Search Related Articles in PubMed Search Related Articles in Pubmed Fair use Excerpt: PubMed Abstract: Bovine inositol polyphosphate 1-phosphatase (1-ptase), M(r) = 44,000, is a Mg(2+)-dependent/Li(+)-sensitive enzyme that catalyzes the hydrolysis of the 1-position phosphate from inositol 1,4-bisphosphate and inositol 1,3,4-trisphosphate. We have determined the crystal structure of recombinant bovine 1-ptase in the presence of ... [ Read More & Search PubMed Abstracts ] end The ID per Robert Smiths' research is most likely a Vaccine - T parva. The surface antigens that he ID'd- 67 kDa- (67p) are a 100% match to the T parva vaccine. One half of the components in the report- are directly related to cows/cattle. Bovine. One half of the components in the report- are related to remediation products- for VOCs - PM 1- which is used for MTBE and other VOCs. The source or vector is likely water and food- beef/milk. Could be biting insects as well. Possibly, cattle vaccinated with the T parva vaccine also consumed water that was remediated - that would explain the source of all the components in Robert Smiths' report. That source explains the disease, the Protozoan/s that Harvey stated they found, the symptoms- the crystals, the microtubles (in T parva) per Darrah (hollow tubes/rods) the skin lesions - T parva causes skin lesions- and much more. It would also explain the presence of Baculovirus- as that is what is in the T parva vaccine. -V- |
|
|
|
Post by Sidney on Sept 16, 2009 13:39:26 GMT -5
|
|
|
|
Post by bannanny on Sept 16, 2009 19:51:10 GMT -5
Wow... that first pic you posted up there kammy looks just like the pattern in this pic of Arni's...  Just comparing... hugs ~~ bannanny |
|
|
|
Post by kammy on Sept 16, 2009 20:39:06 GMT -5
Yes, Banny... I understand that Robert Smith has identified T. parva in a number of people with Morgellons, I'm just not sure what that number is, I'm waiting on Jill to clarify if he's found 100% or what Smith's numbers are? I never found a good link to his work showing his numbers. Like I said, all the researchers should collide eventually and their findings should back each other up if they are correct... at least the main information isn't hidden on the baculovirus and we have a lot to choose from in our reading on this subject. The way I'm seeing it is that most anything could have been put inside or have been incorporated into the bv capsid system within our individual versions, that's why we are all having different symptoms. It is the capsids and their processes that are multiplying at an accelerated pace, they are the main culprit, in that their never ending reproductive processes are keeping us supplied with endless amounts of debris, fibers, spheres, crystals, specks, biofilm, goo and of course, whatever other disease is contained within the bv capsid system is what is creating our individual symptoms. If we eliminate any one component within the 'system', it is not going to necessarily stop it, because it's the baculovirus that's driving the Morgellons Engine that has T. parva, the fungi, the parasites, bacteria, nematodes, the agrobacterium and whatever else is found, as passengers. It's seems that these capsids (cells) are cancer-like in nature, they can reproduce wildly in a very short time and are not controllable by anything that we know of yet. Of course, the scientists have left us some good clues on how to stop them. As an example, if we sought out the remedy for T. parva, or any one component - it's not going to necessarily stop the rest of the entirety of engine's 'system'. We can get a good indication that this is the way it operates by all the different medicines we've taken in the past to attack, let's say two components, such as bacterial and fungal at the same time, and we have seen that approaching our disease this way does not work. It might be because the engine is programmed to regenerate the pathogen at a later time in the life cycle of the system and we have to take into consideration that these pathogens are contained in protective, protein shells or the cell's envelope that are being released at different times. In order to stop the system, we would need to attack the shell, its chemical makeup and the elements that cause it to function and perform its duties that keep the system alive. Jeany and I are studying this bv 'engine' on its core cellular level and trying to figure out what will bring apoptosis or cell death to the engine's system, however, it's complicated, that's why we haven't posted much lately, we're studying. For instance, I was up all last night looking for a protocol for myself and came up with a particular cancer drug along with Interferon, I qualify to take Interferon, however, the cancer drug? I talked with someone today who had taken a different cancer drug with Interferon and it didn't work for them, so, we keep looking. In the meantime, you can see what we're finding out, Jeany has some excellent articles on her blog at morgellons2.wordpress.com/ where she's been putting together some recent information. I'm calling all researchers that have some spare time to help us figure out how this 'system' works and how to stop it?, we can pretty much identify which of the virus types are present from the previous photographs that we've seen and the information about their characteristics. I hope this made sense as to where we're coming from and thank you for bringing this up. Here's a good abstract on apoptosis: jcs.biologists.org/cgi/content/full/117/14/3031 |
|
|
|
Post by fritolay66 on Sept 17, 2009 8:11:11 GMT -5
I hope you don't mind, but I have some thoughts on this. What if it wasn't reproducing at an accelerated rate? Many of us in the past have stated that "we feel as if this has been taking place over the course of a decade". Look at Lyme borrelia. It is a slow, cyclic grower. When it reaches a certain proportion within us, is when one starts to manifest the symptoms in earnest. Even in Bb, not all sufferers exhibit the same symptomology manifestations. The main pathogen is Bb, and then you have the co-infections. They all have the synergistic effect of disease, they all evade or help to evade the immune system. What a perfect way to create human disease and a perfect bio-weapon. I really believe Kammy that the immune system is the key. I don't understand why there aren't more testing results, ie: Morgellons patients under these guidelines. It is my understanding that all the pathogens I have put forth as examples only, are capable of hiding within the macrophages and phagocytes, preventing apoptosis so the cycle can continue. Interferon is a way to increase NK cells. The solution for the many sufferers would be one in which we all would have access to. Raising only the NK cells wouldn't be enough would it? Just my thoughts, thats all. Thanks....  |
|
|
|
Post by Jill on Sept 17, 2009 9:27:17 GMT -5
Kammy and all, You and I agree on certain things- disagree on certain other things.... That Baculovirus is a part of this condition seems certain. What I have read is that Baculovirus- p35- inhibits apoptosis (cell death). That would make sense to me. The components such as the T parva- are kept in check so to speak due to this process. OTH, the normal functions of the body are also inhibited. What I'm saying is: Millions of cells in our bodies die daily- and are re-born or regenerated. This is the normal process of cell death and regeneration. Due to the baculovirus- p35- it seems that this process has been inhibited. Good news- as to the pathogens we are harboring- those pathogens do not continue to regenerate- meaning we don't get worse (at the time). Bad news for us- our bodies are not functioning normally. Could be that all the debris- the specks, fibers and so on are a product of the build up of the cells ?? Our body purges the stuff via lymph - through the skin- outwards... ? Which leads to the question of why the presence of Baculovirus? My theory is that it comes with the rest of the 'finds' by Robert Smith. One half of the components ID'd in his report are related to cattle- COWS. The T parva- source would be a vaccine- the p 67 surface antigen (67 kDa) was ID'd by RS along with the T parva. The fluorescent protein indicates a recombination- ie: man made/lab made. That confirms that it is the vaccine that was ID'd. The vaccine contains that 67 kDa surface antigen- (T Parva) as well as the Baculovirus. ((Other components Robert Smith ID'd are also directly related to COWS- Casein alpha S1- Bovine origin and Leuconostoc mesenteroides subsp. mesenteroides- ATCC 82930 is related to butter/cheese/fermentation processes)) The other half of the components that Robert Smith ID'd are directly related to remediation products- specifically PM 1- to remediate VOC's to include MTBE/TBA/BTEX and pesticides as well. www.thenmo.org/rSmith02.htmtinyurl.com/n2byvvSeems that food and water could be the source?? Milk/milk products/meat- it would seem certain. The Baculovirus that we are dealing with is NOT natural. It is, for certain, lab made. A recombination. jcs.biologists.org/cgi/content/full/117/14/3031Fair use Excerpt: The baculovirus protein P35 inhibits apoptosis in a diverse range of animals such as insects, nematodes and mammals. Evidence suggests that P35 can inhibit members of caspase family proteases that are key mediators of mammalian apoptosis. We demonstrate that p35 inhibits activation-induced nitric oxide (NO)-mediated apoptosis in the RAW 264.7 mouse macrophages. Parent or vector-transfected RAW 264.7 cells underwent apoptosis when treated with a combination of cisplatin and interferon-{gamma} (IFN-{gamma}) or LPS and IFN-{gamma} in a NO-dependent manner.end excerpt Important- most especially the NO or Nitric oxide mentioned (above). Could it be that Nitric oxide will help us? The work of the 6 doctors- Dr Paul cheney, Dr Pall, Dr Garth Nicolson, Dr Teitelman, et all- summarized by Dr Pall: www.prohealth.com/library/showarticle.cfm?id=8071&t=CFIDS_FMBut is it too much Nitric Oxide ? Or not enough. Experts seem to disagree on this important point. Cheney, et al- say to downregulate- Dr Louis Ignarro states we need to supplement with L- Arginine..... www.youtube.com/watch?v=NBPjZJSHr4AAny thoughts on the Nitric oxide issue? Will supplementation help? Or should be use antioxidants to downregulate? Important to me at this time. Thanks for any input, Jill |
|
|
|
Post by kammy on Sept 17, 2009 11:57:11 GMT -5
Thank you Jill, Frito...
They had 6 doctors looking at this, all giving their input to create that report... this is a very complicated system and the more input - the better, it's going to take more than one brain to figure this out, that's for sure. We need help.
We'll be closely looking at each point you've brought up, please ask questions, it helps us all understand better.
From what I've seen under the microscope on how this disease operates, is... there's various capsids or spheres/cells (some which haven't been identified yet) and each have an individual duty or pathogen that it contains. Each of these have been programmed to reproduce specifically in a variety of manners, they are all within a repeating system, this system is centered around the baculovirus delivery processes. (This conclusion is from observation only.)
I think we're making great leaps and bounds into understanding our disease at a faster pace than we have in the past and it's due to our collective research and input as to what's happening and I'm proud of all of us and our various findings!
|
|
|
|
Post by violet on Sept 17, 2009 12:43:33 GMT -5
Thank you Jill, Frito... They had 6 doctors looking at this, all giving their input to create that report... this is a very complicated system and the more input - the better, it's going to take more than one brain to figure this out, that's for sure. We need help. We'll be closely looking at each point you've brought up, please ask questions, it helps us all understand better. From what I've seen under the microscope on how this disease works is... there's various capsids or spheres/cells (some which haven't been identified yet) and each have an individual duty or pathogen that it contains. Each of these have been programmed to reproduce in a variety of manners, they are all within a repeating system, this system is centered around the baculovirus delivery processes. (This conclusion is from observation only.) I think we're making great leaps and bounds into understanding our disease at a faster pace than we have in the past and it's due to our collective research and input as to what's happening and I'm proud of all of us and our various findings! Kammy, thank you again for all you do in your research. I felt so bad for how you were being treated at Morgellons-Disease-Research. Just FYI, I spoke up for you several times there, as I felt the way in which you were spoken to by a couple of the posters was just unbelievably cruel. I just could not believe people would come down on someone like you like they did, as you have always shared information and made it very clear that you hope others will contribute and take part with their thoughts and findings. I'm glad you are here with a more healthy group, Kammy. I don't post there any more, FYI. It's too bad the cuelty continues there towards other people who also do not deserve it. There was a lot of great information shared on that site, but it's just not worth participating in any more. |
|
|
|
Post by kammy on Sept 17, 2009 13:43:14 GMT -5
The Viral Aspect We've heard mention many times of the correlation of a possible herpes involvement in our disease, and wonder what role does it play? If we notice, our disease might be similar to herpes-like and a cancer-like mix? I'll look into this more to see if possibly some of the herpes medicines could help us, to see if the BV is operating similar to herpes? Anyone that wants to help us look, that's great! I found this interesting: Viral genes essential for DNA replication "This set of genes is similar to those identified as being required for herpes simplex 1 transient DNA replication (14) (Table 5.1). In both these viruses it is likely that host enzymes also participate in this process. The similarity of the complement of herpes virus and baculovirus replication and other genes has led to the suggestion that these virus are members of a lineage distinct from other large DNA viruses of eukaryotes." (This table is not showing up properly, see Table 5.1 in the link below:)Table 5.1. Viral genes essential for DNA replication Gene Baculovirus Herpesvirus DNA pol + + (UL30) Ssb + (LEF-3) + (ICP8) primase + (LEF1) + (UL52) Primase accessory factor+ (LEF2) + (UL8) helicase + (p143)+ (UL5) Origin binding protein + IE1 (?) + (UL9) Processivity factor -UL42 Reference: www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=bacvir&part=ch05UL30 - www.wikigenes.org/e/gene/e/2703462.htmlICP8 - www.wikigenes.org/search.html?search=ICP8&adv=0UL52 - www.wikigenes.org/search.html?search=UL52&db=_any&cat=&type=&field=&org=&action=go&ftype=0UL8 - www.wikigenes.org/e/gene/e/2703432.htmlUL9 - www.wikigenes.org/search.html?search=UL9------------------------------------ If you are currently taking l-Arginine or l-Lysine, I would say that it probably can't hurt and might help, until we know more. Here's a web link that shows how the Morgellons lesions can look similar to herpes simplex and some other helpful information: herpessupplements.com/ |
|
|
|
Post by kammy on Sept 17, 2009 14:04:45 GMT -5
Kammy, thank you again for all you do in your research. I felt so bad for how you were being treated at Morgellons-Disease-Research. Just FYI, I spoke up for you several times there, as I felt the way in which you were spoken to by a couple of the posters was just unbelievably cruel. I just could not believe people would come down on someone like you like they did, as you have always shared information and made it very clear that you hope others will contribute and take part with their thoughts and findings. I'm glad you are here with a more healthy group, Kammy. I don't post there any more, FYI. It's too bad the cuelty continues there towards other people who also do not deserve it. There was a lot of great information shared on that site, but it's just not worth participating in any more. Yes, Violet, I have noticed your support throughout and I really appreciate it! People have to realize that we're stumbling into unknown terrains and trying to do the best we can. Of course, we're going to make mistakes, we're supposed to - we're 'laymen' and just figuring it out. This is complicated subject matter and it's going to take us time grasp it and we'll probably never understand everything there is to know. It's not like we're taking someone else's work and using that as a reference here, either... and this fact along with, no scientist out there has come to this same conclusion yet, this puts us in a 'hot seat'. We're blazing a trail for the more experienced to come and help us - is the way I see it. You see how we've all evolved with our expanded knowledges in the various sciences and subjects that we've looked at, we understand through these site forums, more about the way our world and disease is operating? So, if and when somebody comes along that has more expertise, we will be a somewhat-educated panel for them to present our disease to us. We have reason to believe that baculovirus is involved, we're waiting on some collaboration, in the meantime, I see it as - we're trying to educate ourselves. |
|
|
|
Post by fritolay66 on Sept 17, 2009 15:43:07 GMT -5
That kind of behavior will not fly here....  I think you are doing just fine. |
|
|
|
Post by jeany on Sept 17, 2009 15:46:38 GMT -5
After what Fritolay66 has mentioned here that many pathogens hide in the macrophages and phagocytes it made me think about the herpes aspect that might be involved in our disease. It is quite interesting, that many people have reported to have a form of herpes. According to Kammy's research you can see that the Baculovirus and the Herpes virus have similar structural ways to dock on to cells. Herpes viruses also inhibit apoptosis which explains why the symptoms re-occur over and over again. I was thinking that we might need a medication that increases apoptosis in order to eliminate the Baculovirus or at least keep it under control? Something like Aciclovir or Ganciclovir which is used in Herpes treatment? Interesting is also the cancer aspect... www.nature.com/cgt/journal/v7/n4/abs/7700164a.htmlJeany |
|
|
|
Post by kammy on Sept 17, 2009 16:51:12 GMT -5
Apoptosis
To share with you my first search into what might cause apoptosis, I found this first link very informative in a lot of areas of the bv system:www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=bacvir&part=ch03jcs.biologists.org/cgi/content/full/117/14/3031"The baculovirus protein P35 inhibits apoptosis in a diverse range of animals such as insects, nematodes and mammals. Evidence suggests that P35 can inhibit members of caspase family proteases that are key mediators of mammalian apoptosis. We demonstrate that p35 inhibits activation-induced nitric oxide (NO)-mediated apoptosis in the RAW 264.7 mouse macrophages. Parent or vector-transfected RAW 264.7 cells underwent apoptosis when treated with a combination of cisplatin and interferon-{gamma} (IFN-{gamma}) or LPS and IFN-{gamma} in a NO-dependent manner. By contrast, RAW 264.7 cells stably expressing P35 did not undergo apoptosis when treated with a combination of cisplatin and IFN-{gamma} or LPS and IFN-{gamma}. Activation of parent, vector- or p35-transfected cells with cisplatin and IFN-{gamma} or LPS and IFN-{gamma} caused equivalent levels of inducible nitric oxide synthase (iNOS) expression and produced equal amounts of nitrite, which ruled out attenuated iNOS activity during P35-mediated protection. Rather, expression of P35 inhibited translocation of mitochondrial cytochrome c into cytosol, mitochondrial depolarization, activation of caspase-9 and caspase-3, and cleavage of poly (ADP-ribose) polymerase (PARP). These findings indicate that P35 inhibits NO-induced apoptotic cell death of activated macrophages by inhibiting mitochondrial cytochrome c release, which suggests that P35 has targets upstream of the caspase cascade in apoptosis. Nitric oxide (NO) produced by activated macrophages has been implicated in a diverse range of (patho) physiological processes such as cytotoxicity against tumor cells and killing of intracellular pathogens (Nathan, 1992Go; Moncada, 1992Go; Stuehr and Nathan, 1989Go). Macrophages can be activated to produce a large amount of NO by a variety of agents, including cisplatin, a broad-spectrum anticancer drug used for its anti-tumor activity (Son, 1995Go; Sodhi and Suresh, 1992Go). The tumoricidal effect of macrophages has been shown to be further enhanced when these cells are treated with cisplatin in combination with interferon-{gamma} (IFN-{gamma}) (Geetha and Sodhi, 1990Go). The combination of cisplatin-based therapy with an immunostimulant, referred to as chemoimmunotherapy, has shown a synergistic effect in the in vivo antitumor activity of tumor-bearing mice (Johnson et al., 1993Go) and in the in vitro cytotoxic action of immune cells (Pai and Sodhi, 1992Go). Although NO production by activated macrophages is considered to be essential for various tumoricidal and microbicidal functions, macrophages themselves are not immune to the cytotoxic effects of NO. Previously we have reported that mouse peritoneal macrophages treated with a combination of cisplatin and IFN-{gamma} undergo NO-induced apoptotic cell death (Ranjan et al., 1998Go). Others have also provided similar reports on the activation-induced apoptosis of mouse macrophages (Sarih et al., 1993Go; Albina et al., 1993Go). Although various biological effects of NO are known, the molecular mechanism(s) by which NO triggers apoptotic cell death is poorly understood." (**Jill - to answer your question is underlined above...)"Apoptosis is a tightly regulated process of cell death that plays an important role in development, host defense and immune regulation (Borges et al., 2002Go). It is an important biological process for the elimination of unwanted cells such as those with potentially harmful genetic mutation, autoreactive lymphocytes or virally infected cells (Liston et al., 1997Go). Apoptotic events are morphologically distinguishable by cellular shrinkage, nuclear condensation, cytoplasmic vacuolization and membrane blebbing (Wyllie et al., 1980Go). At the biochemical level, apoptosis involves sequential activation of cysteine proteases that cleave after an Asp residue in their substrate, hence also known as caspases (Degterrev et al., 2003). In response to apoptotic stimuli, cytochrome c from the mitochondrial inner membrane space gets released into the cytosol, where it interacts with Apaf-1 to trigger autocatalytic processing of procaspase-9. Caspase-9 then activates caspase-3 and other effecter caspases, resulting in the proteolytic cleavage of substrate nuclear poly(ADP-ribose)polymerase (PARP) (Fesik, 2000Go). Importantly, PARP cleavage has been observed in a variety of apoptotic responses, including oxidative stress-mediated cell death (Tewari et al., 1995Go). Direct evidence for the involvement of caspase family proteases in apoptosis comes from studies using the cowpox virus protein CrmA and the baculovirus protein P35, which are direct inhibitors of at least certain members of this enzyme family (Clem and Miller, 1994Go). To circumvent host defense mechanisms, viruses have evolved mechanisms to antagonize host death signals so that viral propagation can continue unabated in infected cells (Oltvai and Korsmeyer, 1994Go). Baculovirus protein P35 has been shown to inhibit virally induced apoptosis, developmental cell death in Caenorhabditis and Drosophila, and neuronal cell death in mammals (Clem and Miller, 1994Go; Rabizadeh et al., 1993Go). Further, it has been shown to confer protection against various inducers of apoptosis including oxidative stress, TNF-{alpha} and Fas (Tewari et al., 1995Go). These reports suggest that P35 must interrupt a highly conserved and ubiquitous component of the death machinery. Recent findings suggest that P35 prevents cell death by inhibiting enzyme activity of several members of the caspase family proteases, including mouse caspase-1, human caspase-1 and human CPP32, by forming stable complex, thereby competitively inhibiting these proteases (dela Cruz et al., 2001Go). Despite the present understanding of P35-mediated inhibition of apoptosis, the precise mechanism involved therein is unclear. In the present study, we investigated the mechanism of inhibition of NO-induced apoptosis in cisplatin and IFN-{gamma}-, or LPS and IFN-{gamma}-activated RAW 264.7 macrophages by baculovirus P35 and its potential targets." ------------------------------------- This above took me to look closer at cisplatin:en.wikipedia.org/wiki/CisplatinCisplatin is an anti-neoplastic agent ------------------------------------- Looking closer at what is an anti-neoplastic agent:en.wikipedia.org/wiki/Alkylating_antineoplastic_agent------------------------------------- Here is study done on mice that indicates that MMS (this is NOT the same as the drops that we've been taking), this is a cancer drug called Methyl methanesulfonate along with cyclophosphamide, another cancer drug - in which they were compared to each other along with drinking Orange Juice. The MMS preformed better than the CP.
How this might help someone?: if you're currently taking a cancer drug and it's not helping with your "M" symptoms, you might ask them to try the MMS, instead? And, by us drinking more orange juice alone, might help with some of our symptoms?mutage.oxfordjournals.org/cgi/content/full/20/4/279"Influence of orange juice over the genotoxicity induced by alkylating agents: an in vivo analysis There is considerable epidemiological evidence indicating an association between diets rich in fresh fruit and vegetables and a decreased incidence of cancers. Methyl methanesulfonate (MMS) and cyclophosphamide (CP) are alkylating agents that differ in their mode of action. MMS is a directly-acting, monofunctional agent, while CP is a bifunctional agent that requires metabolic activation to a reactive metabolite. To evaluate if orange juice could reduce DNA damage induced by these alkylating agents, mice were treated orally (by gavage) with MMS and CP, prior to and after treatment with orange juice. DNA damage was evaluated by the comet assay in peripheral white blood cells. Under these experimental conditions, orange juice reduced the extent of DNA damage caused by both mutagens. For MMS, the antigenotoxic effect of the orange juice was both protective (orange juice pre-treatment) and reparative (orange juice post-treatment); for CP, the effect was reparative only. The components of orange juice can have several biological effects, including acting as targets of toxicants and modulating metabolization/detoxification routes. Considering the different mechanisms of the action of the two drugs, different protective effects are suggested. These results demonstated the ability of the in vivo comet assay to detect in vivo modulation of MMS and CP mutagenicity by orange juice. Diet represents a major influence on the promotion and progression of cancer. A micronutrient-equilibrated diet can contribute to genomic stability. Deficiencies in vitamins and minerals in the human diet are thought to generate DNA damage by enhancing the occurrence of breaks and oxidative lesions (1GoGo–3Go). Since mutations are key elements in neoplasic processes, there is a considerable amount of epidemiological evidence relating diets rich in fresh fruit and vegetables and a decrease in cancer incidence (2Go,4Go)." -------------------------------- Cyclophosphamide or 'CP' en.wikipedia.org/wiki/Cyclophosphamide"Cyclophosphamide is a potent antitumour agent used against many forms of cancer and against certain other diseases. Chlorophyllin, which is obtained by hydrolysis of chlorophyll to remove phytyl alcohol, is an efficient antimutagenic agent and has been used as a dietary supplement or to diminish the intensity of the discomforting side effects of cyclophosphamide therapy. Chlorophyll may have beneficial effects when used in combination with cyclophosphamide therapy." We see the benefits of using chlorophyll or chollera, which has been brought up in the past, as possible benefits with our treatments. |
|
|
|
Post by jeany on Sept 17, 2009 16:57:13 GMT -5
This is from my blog: morgellons2.wordpress.com/2009/09/07/insect-virus-proteins-falpe-and-p10-self-associate-to-form-filaments-in-infected-cells/Insect virus proteins (FALPE and p10) self-associate to form filaments in infected cells
The main symptom of Morgellons Disease is the appearance of fibers emerging from intact skin or in other cases emerging from open sores. All, namely 100%, Morgellons Sufferers report this condition. According to scientists investigating this disease it has not been proven to this day what causes the formation of these fibers. The exact chemical structure of the fibers is not fully understood nor investigated and still remains unknown. This article will show how the Entomopoxvirus, and the Baculovirus, both used in combination as a bio-insecticide, is kept suspiciously responsible for the cause of this life threatening disease. Here are a few excerpts from this article which I’ve found quite interesting: Entomopoxviruses and baculoviruses are pathogens of insects which replicate in the cytoplasm and nuclei of their host cells. During the late stages of infection, both groups of viruses produce occlusion bodies which serve to protect virions from the external environment. * We remember that occlusion bodies are also part of the Baculovirus. Description Occlusion Bodies: www.uniprot.org/keywords/842Baculoviruses and Entomopoxviruses belong to the family of Cypoviruses. continuing excerpt: Immunofluorescence and electron microscopy studies have shown that large bundles of filaments are associated with these occlusion bodies. Entomopoxviruses produce cytoplasmic fibrils which appear to be composed of the filament-associated late protein of entomopoxviruses (FALPE). Cytoplasmic fibrils  Evidence that FALPE and p10 could produce filaments in the absence of other viral proteins is presented. When FALPE was expressed in insect cells from a recombinant baculovirus, filaments similar to those produced by the wild-type Amsacta moorei entomopoxvirus were observed.
In addition, when expression plasmids containing FALPE or p10 genes were transfected into Vero monkey kidney cells, filament structures similar to those found in infected insect cells were produced.The manner in which FALPE and p10 subunits interact to form polymers was investigated through deletion and site-specific mutagenesis in conjunction with immunofluorescence microscopy, yeast two-hybrid protein interaction analysis, and chemical cross-linking of adjacent molecules. * As we know, scientists investigating Morgellons Disease claim to have found that the related fibers contain a source of polymer and also fungi/yeast plays a significant role. source: www.ncbi.nlm.nih.gov/pubmed/9499079Another article according to the protein p10: source: www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=bacvir&part=ch02Fibrous p10-containing material aligned  excerpt: P10 (Ac137). Although p10 does not appear to be a major occlusion body protein, it colocalizes with the PE protein and appears to be required for the proper formation of the polyhedron envelope. When p10 is phosphorylated, it becomes associated with microtubules (15). This could be related to the structures it forms that include microtubule-associated filaments, and tube-like structures that surround the nuclei of infected cells. Us, Morgellons Sufferers, have discussed the effects of Global Warming and its effects on nature. We are assuming that this condition is responsible for the enhancement and over population of insects causing this disease. jvi.asm.org/cgi/content/full/72/3/2213?maxtoshow=&HITS=10&hits=10&RESULTFORMAT=&fulltext=mating&searchid=1&FIRSTINDEX=1390&resourcetype=HWFIGEntomopoxviruses and baculoviruses are pathogens of insects which replicate in the cytoplasm and nuclei of their host cells, respectively. During the late stages of infection, both groups of viruses produce occlusion bodies which serve to protect virions from the external environment.
Immunofluorescence and electron microscopy studies have shown that large bundles of filaments are associated with these occlusion bodies.
Entomopoxviruses produce cytoplasmic fibrils which appear to be composed of the filament-associated late protein of entomopoxviruses (FALPE). Baculoviruses, on the other hand, yield filaments in the nuclei and cytoplasm of the infected cell which are composed of a protein called p10.
Evidence that FALPE and p10 could produce filaments in the absence of other viral proteins is presented. When FALPE was expressed in insect cells from a recombinant baculovirus, filaments similar to those produced by the wild-type Amsacta moorei entomopoxvirus were observed. In addition, when expression plasmids containing FALPE or p10 genes were transfected into Vero monkey kidney cells, filament structures similar to those found in infected insect cells were produced. The manner in which FALPE and p10 subunits interact to form polymers was investigated through deletion and site-specific mutagenesis in conjunction with immunofluorescence microscopy, yeast two-hybrid protein interaction analysis, and chemical cross-linking of adjacent molecules. These studies indicated that the amino termini of FALPE and p10 were essential for subunit interaction. Although deletion of the carboxy termini did not affect this interaction, it did inhibit filament formation. In addition, modification of several potential sites for phosphorylation also abolished filament assembly. We concluded that although the sequences of FALPE and p10 were different, the structural and functional properties of the two polypeptides appeared to be similar. Cytoskeletal elements have previously been demonstrated to be involved in several aspects of virus assembly. For example, vaccinia virus has been shown to associate with actin during its release from the plasma membrane , while adenovirus is transported through the cytoplasm to the nucleus through its interaction with microtubules. Actin has been implicated in the transport of baculovirus nucleocapsids to the nucleus. Other viruses contain actin in their envelopes along with viral surface glycoproteins, implying some role in the budding process. In addition, cytochalasin D, a disruptor of microfilaments, has been shown to impair the assembly of a number of different viruses. Most viruses use preexisting microtubule or microfilament proteins derived from host cells in these processes. However, we have recently demonstrated that insect poxviruses establish their own filament network during the later stages of infection, using a protein encoded by the viral genome. AmEPV derives its name from the Indian red army worm, a larva from the Lepidoptera family and the host from which the virus was originally isolated. Lepidoptera Baculoviruses also infect Lepidoptera larvae but instead replicate in the nuclei of their host cells . A number of baculoviruses have been studied, but knowledge of Autographa californica nuclear polyhedrosis virus (AcNPV), which infects a wide variety of larvae including that of the alfalfa leaf hopper, is most extensive. This virus is used routinely to produce recombinant proteins in insect virus expression systems. A common property of EPVs and baculoviruses is the formation of large intracellular structures known as occlusion bodies which assemble during the late stages of viral infection. Virions are embedded within these occlusion bodies, and the process serves to protect the virus from the external environment. In the case of baculoviruses, the occlusion bodies are called polyhedra and are composed predominantly of a 31-kDa protein called polyhedrin . The occlusion bodies of EPVs are known as spheroids and consist mainly of a 110-kDa protein known as spheroidin. Spheroidin and polyhedrin do not appear to exhibit sequence homology. A multilamellar envelope also appears to surround both polyhedra and spheroids and may help to stabilize these structures during assembly. During the late phases of AmEPV and baculovirus infections, large bundles of filaments also appear to accumulate in the infected insect cells.In the case of AmEPV, these structures are present in the cytoplasm, while those found in cells infected with baculoviruses reside both in the cytoplasm and in the nucleus . Baculovirus fibrils are composed primarily of a 10-kDa protein called p10. Deletion mutagenesis of AcNPV p10 has demonstrated that both the amino- and carboxy-terminal regions of this protein are necessary for the formation of filaments in the infected cell. Our laboratory recently demonstrated that the cytoplasmic filaments, which characterize the late stages of infection by AmEPV, are composed primarily of a 156-amino-acid protein called FALPE (filament-associated late protein of EPVs). These filaments are closely associated with the spheroids and their membrane envelopes. FALPE is a phosphoprotein which migrates on sodium dodecyl sulfate (SDS)-polyacrylamide gels as a 25/27-kDa doublet. This protein also contains an unusual proline-glutamic acid repeat region spanning 20 residues in the carboxy terminus of the polypeptide. The ultrastructure and close association of this protein with the occlusion bodies of AmEPV suggested that FALPE and p10 played analogous roles during infections by the respective viruses. This article addresses the structural and functional similarities between FALPE and p10. These two viral proteins are known to be major components of filamentous structures, but it is not known whether additional viral or cellular proteins cooperate during the polymerization process. In this report, we provide insight into the mechanisms which produce filaments in cells infected with either baculoviruses or EPVs. We demonstrate that p10 and FALPE can produce filaments in the absence of other viral gene products. Using the yeast two-hybrid system and a chemical cross-linking agent, we obtained evidence for self-association of either FALPE or p10. Finally, the polypeptide regions of FALPE and p10 which are required for self-association and subsequent filament formation are mapped.  showing filaments associated with FALPE in Sf9 insect cells infected with AmEPV and a FALPE recombinant baculovirus. Sf9 cells were mock infected (A), infected with AmEPV (B), infected with wild-type AcNPV (C), or inoculated with a recombinant AcNPV expressing the FALPE gene (D). At 72 h postinfection, cells were incubated with MAb CLP001, and bound antibody was detected with goat antimouse antibody conjugated to fluorescein. Labeled proteins were visualized with a Leitz fluorescence microscope. Nuclear DNA in panels A and D was also stained with Hoescht dye, and the two panels represent double exposures from the fluorescein and Hoescht signals. Panels B and D show the filaments formed by FALPE when expressed by AmEPV and AcNPV, respectively, while panel C illustrates background fluorescence found in the nuclei of cells infected with wild-type AcNPV.  Immunofluorescence microscopy of Vero monkey kidney cells transfected with a FALPE or p10 expression plasmid. Vero cells were transiently transfected with either the expression plasmid pRBK (A), plasmids expressing the p10 gene of AcNPV (B and D), or a plasmid expressing FALPE (C). FALPE was visualized by using MAb CLP001 (C), while p10 was detected with a rabbit polyclonal antibody directed against the p10 protein of AcNPV (B and D). Control cells in panel A were stained with the p10 polyclonal antibody; nuclei in panel D were also stained with Hoescht dye. Clearly both p10 and FALPE formed filament networks following transient transfections of their genes into mammalian cells.  Immunofluorescence microscopy showing intracellular localization of FALPE mutant proteins produced by recombinant baculoviruses. Sf9 insect cells were infected with baculoviruses expressing N45.FALPE (A), 8098.FALPE (B), Ct.FALPE (C), or the phosphorylation mutant P Mut.FALPE (D) as described for Fig. 2. MAb CLP001 and fluorescein isothiocyanate-coupled goat anti-mouse antibody were used to detect FALPE variants in panels A, B, and D. The mutant Ct.FALPE was detected by using a rabbit polyclonal primary antibody directed against FALPE and rhodamine-coupled goat anti-rabbit secondary antibody. Nuclear DNA in panels A and B was also stained with Hoescht dye, and the photographs represent double exposures of fluorescein and DNA signals. All mutations abolished the ability of FALPE to produce filaments in cells infected with the different recombinant baculoviruses. The bars in panels A and C represent 10 µm, while those in panels B and D indicate 15 µm. Jeany |
|
|
|
Post by jeany on Sept 17, 2009 17:03:16 GMT -5
morgellons2.wordpress.com/2009/09/12/formation-of-actin-filaments-in-mammalian-cells-baculovirus-protein-falpe-and-p10/Formation of Actin Filaments in Mammalian Cells – Baculovirus Protein FALPE and p10
FALPE and p10 Insect virus protein embedded in the Baculovirus might cause the proliferating formation of Actin Protein Filaments in humans bodies..mainly Morgellons Sufferers. As we know, the so called ‘fibers’ are seen to be the main symptom of this disease. 100% have reported to have them in/on lesions or have them emerging from intact skin or hair follicles. I believe that the infectious Baculovirus leads to an over reaction of Actin Protein filament production in our organism due to influence of the Insect virus proteins FALPE and p10. Here are a few excerpts out of the explanation of Actin Protein Filaments: Actin is a globular, roughly 42-kDa protein found in all eukaryotic cells. Actin is the monomeric subunit of two types of filaments in cells: microfilaments, one of the three major components of the cytoskeleton, and thin filaments, part of the contractile apparatus in muscle cells. Thus, actin participates in many important cellular processes including muscle contraction, cell motility, cell division and cytokinesis, vesicle and organelle movement, cell signaling, and the establishment and maintenance of cell junctions and cell shape. Principal interactions of structural proteins are at cadherin-based adherens junction. Actin filaments are linked to α-actinin and to the membrane through vinculin. The head domain of vinculin associates to E-cadherin via α-, β-, and γ-catenins. The tail domain of vinculin binds to membrane lipids and to actin filaments. The protein actin is one of the most highly conserved throughout evolution because it interacts with a large number of other proteins. All non-spherical prokaryotes appear to possess genes such as MreB, which encode homologues of actin; these genes are required for the cell's shape to be maintained. The plasmid-derived gene ParM encodes an actin-like protein whose polymerised form is dynamically unstable, and appears to partition the plasmid DNA into the daughter cells during cell division by a mechanism analogous to that employed by microtubules in eukaryotic mitosis. Actin has four main functions in cells : To form the most dynamic one of the three subclasses of the cytoskeleton, which gives mechanical support to cells, and hardwires the cytoplasm with the surroundings to support signal transduction. To allow cell motility (see Actoclampin molecular motors), including phagocytosis of bacteria by macrophages. In muscle cells to be the scaffold on which myosin proteins generate force to support muscle contraction. In non-muscle cells as a track for cargo transport myosins [non-conventional myosins] such as myosin V and VI. Non-conventional myosins transport cargo, such as vesicles and organelles, in a directed fashion, using ATP hydrolysis, at a rate much faster than diffusion. Myosin V walks towards the barbed end of actin filaments, while myosin VI walks toward the pointed end. Most actin filaments are arranged with the barbed end toward the cellular membrane and the pointed end toward the cellular interior. This arrangement allows myosin V to be an effective motor for export of cargos, and myosin VI to be an effective motor for import. Pathway of Baculovirus in mammalian cellsWe have studied the infection pathway of Autographa californica multinuclear polyhedrosis virus (baculovirus) in mammalian cells. By titration with a baculovirus containing a green fluorescent protein cassette, we found that several, but not all, mammalian cell types can be infected efficiently. We demonstrate for the first time that this baculovirus can infect nondividing mammalian cells, which implies that the baculovirus is able to transport its genome across the nuclear membrane of mammalian cells. Our data further show that the virus enters via endocytosis, followed by an acid-induced fusion event, which releases the nucleocapsid into the cytoplasm.
Cytochalasin D strongly reduces the infection efficiency but not the delivery of nucleocapsids to the cytoplasm, suggesting involvement of actin filaments in cytoplasmic transport of the capsids.In one study, nucleocapsids of a related baculovirus species (Ploidia interpunctella granulosis virus) were observed docking at the nuclear pore of infected insect cells, at different stages of releasing their genome, but not inside the nucleus. This suggests a mechanism of DNA transport similar to HSV.We show here that baculovirus can indeed infect nondividing mammalian cells through a mechanism apparently identical to that found in insect cells. The mode of nuclear entry of the viral genome appears to be different from what is known of other large DNA viruses. Our data suggest that the cigar-shaped nucleocapsid (25 nm in diameter) is transported through the nuclear pore, together with the viral genome. References: en.wikipedia.org/wiki/Actinwww.pubmedcentral.nih.gov/articlerender.fcgi?artid=113992Jeany
|
|
|
|
Post by kammy on Sept 17, 2009 17:14:26 GMT -5
Wow... that first pic you posted up there kammy looks just like the pattern in this pic of Arni's... Just comparing... hugs ~~ bannanny It is an interesting comparison, Banny. We are wondering about these 'fish' looking objects... it used to be the 'seashells' that I thought I'd never understand what they meant... now, it's the 'fish'?... this is one weird disease, that much, I know. Those are very informative articles, Jeany, thank you for sharing. |
|
|
|
Post by kammy on Sept 17, 2009 17:23:41 GMT -5
The Lymes / Morgellons Link We know that Morgellons is a co-infection of Lymes, and Lymes is a co-infection of Morgellons. So, what do they have in common? Most likely an insect? The Tick = T. parva? Has this particular tick or the cow vaccine containing T. parva been linked to Lymes? Maybe Jill can tell us how many of the Lymes co-infections are being found by Robert Smith to also be a part of Morgellons, or has he concentrated in that area? Or, at what numbers Bb or Wolbachia is being found with "M" samples? Anyone have any data on this? I'm for certain that I have T. parva as a part of my Morgellons, I had someone ask me today about these 'cones'... and told them that Jill could better answer this since she is following Smith's findings closely. What is T. parva's role in our disease? Jill, do you have info on the 'cone', 'glue', 'cement', aspects of the lesions? We see at the end of page 6, where I question GP64's role, which we know to be present in human samples, which contains a tick element of the Thogotovirus (family Orthomyxoviridae), which are pathogenic for ticks and can also infect vertebrates.: What family is T. parva in? Is this our relationship to Lymes? www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=11991958In earlier investigations of baculovirus biology, a protein called GP64 was identified as the envelope fusion protein of AcMNPV and its close relatives (2, 3, 42, 47). Subsequently, it was discovered that GP64 is closely related (∼30% identity at the amino acid level) to the envelope fusion protein encoded by members of the genus en.wikipedia.org/wiki/Thogotovirus Thogotovirus (family Orthomyxoviridae), which are pathogenic for ticks and can also infect vertebrates (22, 27, 29, 33). This indicated that an envelope fusion protein either had been transferred between a minus-sense RNA virus and a double-stranded DNA virus or was acquired independently by each of these viruses from another source, such as a conserved host gene (29) (Fig. ​(Fig.11FIG. 1.) Theoretically speaking, I am stating that from what I have presented that people with Lymes without Morgellons do not have the baculoviral aspect as part of their disease, because the symptoms of BV are specks, biofilm and fibers. The people with Lymes and Morgellons that have these symptoms, do have baculoviral involvement. |
|
|
|
Post by Jill on Sept 17, 2009 19:57:38 GMT -5
The Lymes / Morgellons Link Has this particular tick or the cow vaccine containing T. parva been linked to Lymes? ((Yes- It is linked to Lyme disease)) Maybe Jill can tell us how many of the Lymes co-infections are being found by Robert Smith to also be a part of Morgellons, or has he concentrated in that area? Or, at what numbers Bb or Wolbachia is being found with "M" samples? Anyone have any data on this? ((see below)) I'm for certain that I have T. parva as a part of my Morgellons, I had someone ask me today about these 'cones'... and told them that Jill could better answer this since she is following Smith's findings closely. What is T. parva's role in our disease? Jill, do you have info on the 'cone', 'glue', 'cement', aspects of the lesions? . Kammy and all, I can not answer how many of the Lyme co-infections were found by Robert Smith- that are a part of Morgellons- just that 1/2 of the components relate to cattle -T parva, Casein alpha S1- Bovine origin and Leuconostoc mesenteroides subsp. mesenteroides- ATCC 82930 is related to butter/cheese/fermentation processes. The other 1/2 of the components- relate to bacterias/components in remediation (water/soil cleaning) products- primarily PM 1- which remediates VOCs - MTBE/TBA/BTEX and pesticides. The Nocardiodes RS ID'd, in addition to being used in wastewater treatment (remediation), is a source of DEEP FUNGAL INFECTIONS. There are too many variables to list with just what RS reported. It does tie- beyond a shadow of a doubt to what we are experiencing- and it is found in the environment. The Rhipicephalus appendiculatus-TICK is the vector of T parva, however, I believe that the T parva that RS ID'd is sourced from a vaccine- most likely vectored via the food chain (meat/milk/cheese)- from cattle vaccinated with the T parva 67 kDa surface antigen- which is the contagious portion the sporozoites, which enter lymphocytes and form the Schizonts. Other studies show that many pathogens are found in the same sample (sufferer/victim). One Russian study I was reading today, stated that as many as 11 or 12 are found in the same sample. That could be Bb, Babs, T parva and so on. Just as we have co-infections of Lyme disease, it seems that there are numerous possible co-infections of T parva. In Africa, Babesia is common. Back to the cones- the tissue cement patent explains the cone and the cortex (center) of the cone that is formed by the tick. Attached to the hosts' skin: United States Patent: 7335733 This cement forms a bond that is better than that of anything on the market: Fair use Tissue cement excerpt: Such a pharmaceutical composition has many applications, particularly in skin surgery and wound healing, for the temporary or permanent bonding of human or animal tissues to each other or to other biomaterials. Tissue cement has previously been used in surgical procedures to provide adhesion and stability to living tissues to enable the normal processes of healing and repair to take place or to provide a long term bond in situations where normal healing is delayed or unlikely to occur Tissue cement formed from the proteins of the present invention have several advantages over conventional tissue cements For example, the proteins of the invention form strong bonds with vertebrate tissues. This makes them ideal for use as components of a tissue cement to bond two tissue surfaces or edges together end More on the cones/T parva later- |
|